DNA has come to occupy a central place in the scientific and popular imaginary as an arbiter of identity and a determiner of biological outcomes. However the assumptions we make about DNA’s centrality rely on a series of underestimates and negations.
DNA is often presented as the holder of the ‘code.’ The code, broadly speaking, is understood as a set of biological instructions, sent downstream within the cell, that determine how the cell behaves and how the organism develops and functions (with, of course, some input from ‘the environment’).
In this understanding, the significance of DNA’s networked interactions with other biomolecular species and processes is vastly understated, as many biologists and philosophers of biology have argued (such as Michel Morange, Susan Oyama). DNA acts on, and is itself acted upon, by an unfathomably complex, networked series of cellular processes, with biological outcomes emerging from the intricate relationality of these collaborative forces. These forces include the genes themselves and their intricate interactions and feedback loops, and significantly, extra-genomic processes such as: epigenetic processes that produce heritable modifications to gene expression without changing the genetic code itself; variable post-transcriptional processing of mRNA; and alternative 3-dimensional shapes protein molecules can fold into, which radically changes their function. These processes are all influenced by constantly changing conditions within the cell, moment to moment. Broader forces such as behavioural, cultural and environmental processes are also integral actors in this cellular conversation.
Seen from an entirely different perspective, we also underestimate DNA’s biological messiness. In our conception of DNA as ‘code’ we imagine DNA to be neatly ordered and packaged into the nuclei of cells (with some extra DNA neatly packaged into mitochondria) where it acts as a logical code, issuing instructions to the rest of the cell. This DNA imaginary fails to acknowledge the ways in which DNA exceeds these bounds and roams rogue beyond the cell: mingling, exchanging, drifting, persisting beyond known time, and acting as an agential force in otherwise unexpected ways.
My broad goal in this residency was to ‘denature’ DNA by asking: How does DNA exceed the spatial, material, temporal and ethical bounds we place around it in our habitual understandings? How does DNA go rogue? When is DNA ‘out-of-place’?
My practice more broadly explores philosophically and experientially ambiguous zones in relation to life. Previous projects have explored the life-death boundary, the intersubjective nature of organ transplantation, and the animate-inanimate boundary in relation to prosthetics. I use living entities such as cells and organs, and in my most recent work, my own surgically excised bone material, to devise conceptual, aesthetic and experiential strategies that bring liminal or unacknowledged processes into more conscious awareness.

My previous project The End is a Distant Memory (2016) and collaborative project with Peta Clancy The Body is a Big Place (2011-2013) explored the capacity for organs, cells, and biomolecules to retain function long after death of the whole organism. This revealed death as a durational process, rather than an event that takes place in a moment of time. This complexity generates enormous bioethical ambiguity for contemporary medical practices that rely on precise determination of death, for example organ retrieval from cadaveric donors for organ transplantation. The residency used this prior work as a departure point to think about liminality and DNA.

The focus of the proposed residency research was on 2 overlapping areas:
DNA liminality
DNA ownership and governance
DNA liminality
I was interested in researching the liminality of donated DNA material. Drawing on the expertise of Jimmy and Caroline I hoped to understand the legal, ethical, governance and ownership status of DNA material as it moves through increasing levels of abstraction, from patient body to excised wet tissue samples, to sequenced genetic code, to silicon-based algorithmic code, to large-scale complex genomic data sets and infrastructure, during the process of biomedical research. This represents a degree of biological abstraction I had not yet explored in my practice.
I was keen to explore the ontological shifts that take place as donated DNA moves through these increasing levels of abstraction. Biomaterial that leaves our body retains biological signifiers of our identity, and can retain the potential for biological function for some time. Can these DNA samples still be regarded as ‘living material’ and at what point is this ‘aliveness’ no longer present? At what point does an individual lose personal investment in their DNA material? These questions were of particular interest to me due to DNA’s iconic position in the scientific and public imagination.
As the residency developed these questions expanded significantly into new areas I hadn’t considered before, which are described elsewhere in this blog.
DNA ownership and governance
In 2019 I had hip replacement surgery, required due to a congenital hip abnormality in my right hip. I hoped to obtain the femur bone removed during my surgery to make a bone china object, to create a quasi-ritualistic process to honour the emotional, symbolic and mythic investments I had in my bone, and to explore adaptation to bodily change, part of a work called Habitation.
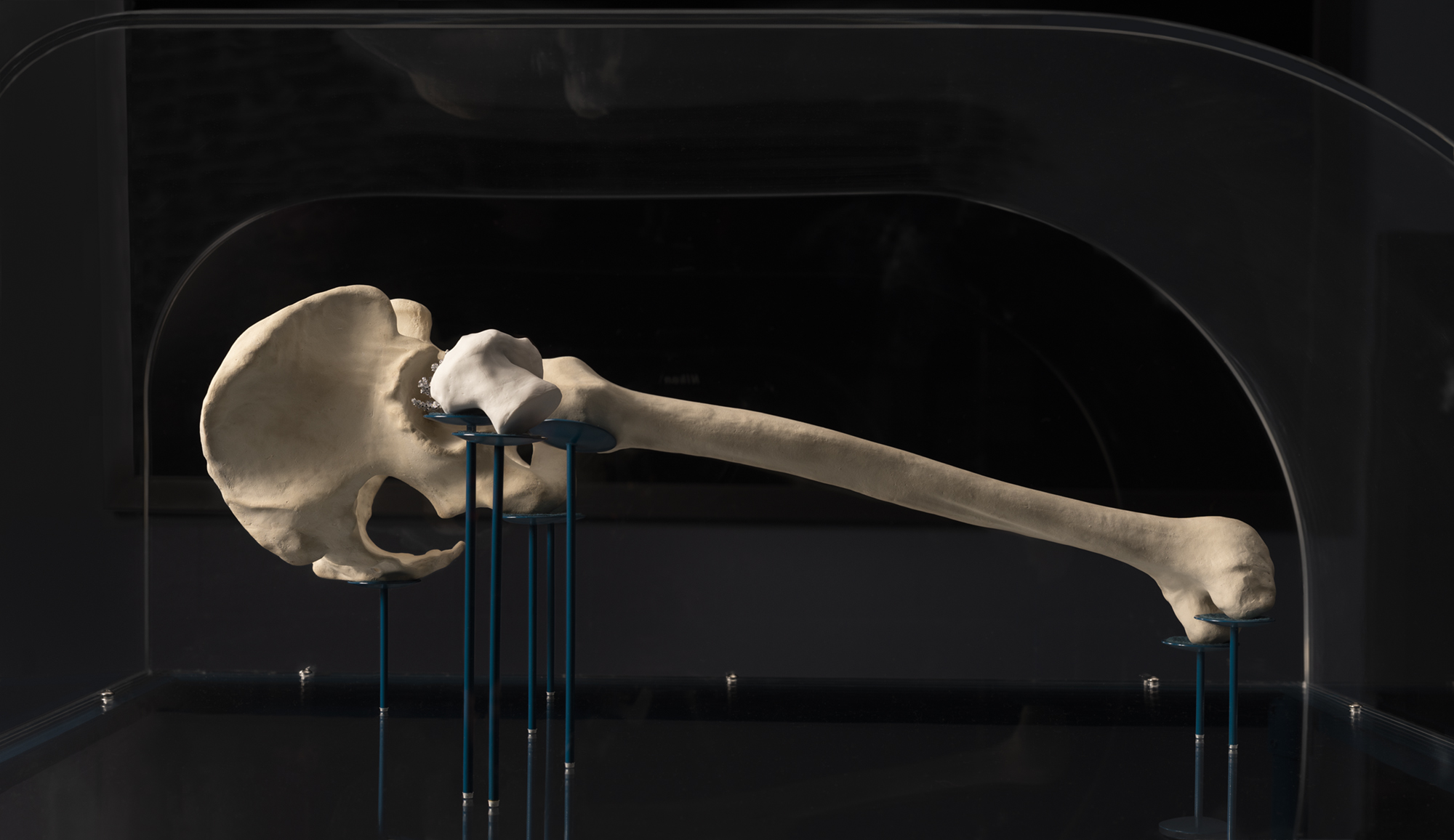
The process of gaining permission to keep my bone was not straight forward and relied on research my colleague Jaden J. A. Hastings had undertaken to obtain her own surgically excised tissues. The policies of the Federal Department of Health and the Royal Australian College of Pathologists, in relation to ‘patient retention’ of surgically excised tissues, unambiguously support patients to retain their excised biomaterial if desired, unless there are specific counter arguments against it. These arguments can include public health risks the tissue poses, or if the tissue has been substantially altered after surgical excision, for example through pathology testing, intellectual property rights are conferred to the pathology company or entity who undertook the modifications. Surgeons, and indeed my own surgeon, are frequently unaware that the default policies support patient access to their own biomaterials.
The lack of a shared understanding between the community, policy makers, legal and medical practitioners about the status of excised human biomaterial underscores the ambiguous, liminal space human tissue occupies once it leaves the originating body. This may reveal unspoken, sometimes conflicting assumptions and beliefs held by individuals, institutions and corporations about excised human biomaterial.
I was excited that Carolyn would be part of the residency collaboration as I hoped she could guide me to a clearer understanding of how legal frameworks grapple with the inherent liminality of biological entities, and whether this might make apparent some of these unspoken assumptions and contradictions.
Another aspect of the proposed residency research concerns DNA donors. Jimmy and his research group work with DNA material donated by leukemia and other patient groups, which is sequenced for the purposes of research. Jimmy notes that DNA donors don’t fully appreciate the significant value and legacy of their DNA material, through its use in genetic research and the potential development of new treatments and medical innovations.
An intention of the residency research was to explore conceptual, representational and experiential strategies to both highlight the liminality of donated DNA, and make more tangible its material, codified and knowledge-generating existences, to those outside the biomedical research community. Jimmy and I hoped to devise speculative approaches for restoring some of the lost connectivity between DNA donors and their DNA material. Jimmy explained that this can be a complex issue for some patient groups such as cancer patients, who may have ambivalent attitudes to the DNA that has ‘betrayed’ them.
Creative work
The residency was structured as a research residency and when I started the residency I had no firm plans for specific artworks, but I hoped new works would emerge from the research. The research has spawned a new work-in-development exploring a particular instance of DNA’s ‘excess,’ which is detailed elsewhere in the blog.
A word on genomic hegemony and post-genomic biology
In reference to what I wrote at the beginning of this post … As early as 1957 American geneticist, cell and developmental biologist David L. Nanney articulated what’s come to be known as the genomic hegemony, and its alternative which he terms the ‘Steady State’ model but which today we might call post-genomic biology. He frames these tensions graphically and metaphorically in overtly political terms. Our understanding of the relational nature of genetic and cellular processes has developed enormously since 1957 but DNA’s hegemonic grip has not loosened easily, at least in part due to the cultural, financial and ideological investments embedded within these ideas, as Susan Oyama, Evelyn Fox Keller and other philosophers and sociologists of biology have explored.
Nanney writes:
Two concepts of genetic mechanisms have persisted side by side … The first of these we will designate the ‘Master Molecule’ concept … In its simplest form the concept places the ‘master molecules’ in the chromosomes and attributes the characteristics of an organism to their specific construction; all other cellular constituents are considered relatively inconsequential except as obedient servants of the master. This is in essence the Theory of the Gene, interpreted to suggest totalitarian government…
The second concept of a genetic mechanism is one which is more difficult to describe … This concept we will designate as the ‘Steady State’ concept. By the term ‘Steady State’ we envision a dynamic self-perpetuating organization of a variety of molecular species which owes its specific properties not to the characteristics of any one kind of molecule, but to the functional interrelationships of these molecular species… In contrast to the totalitarian government by ‘master molecules,’ the ‘steady state’ government is a more democratic organization, composed of interacting cellular fractions operating in self-perpetuating patterns.
D. L. Nanney, “The Role of the Cytoplasm in Heredity.” In The Chemical Basis for Heredity, eds W.D. McElroy and B. Glass (Baltimore, Maryland: John Hopkins University Press, 1957), 134; quoted in D. Haig, “The (Dual) Origin of Epigenetics.” Cold Spring Harbour Symposia on Quantitative Biology, Volume LXIX. (Cold Spring Harbour, NY: Cold Spring Harbour Laboratory Press, 2004): 68.